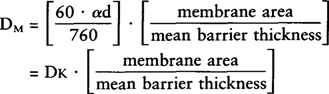
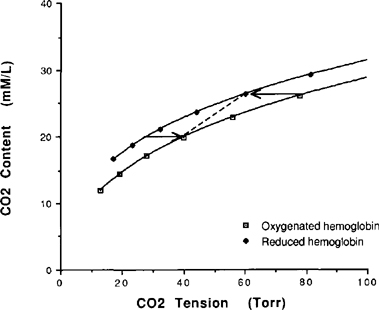
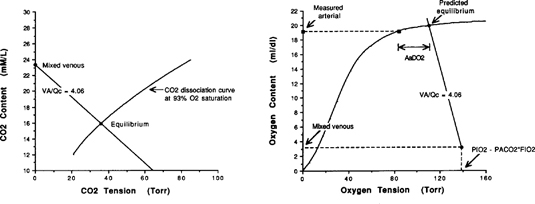
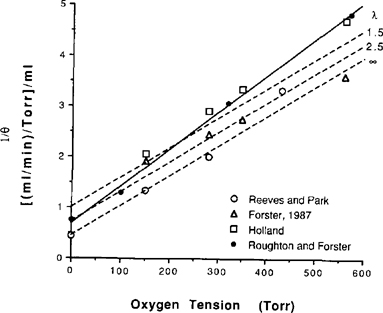
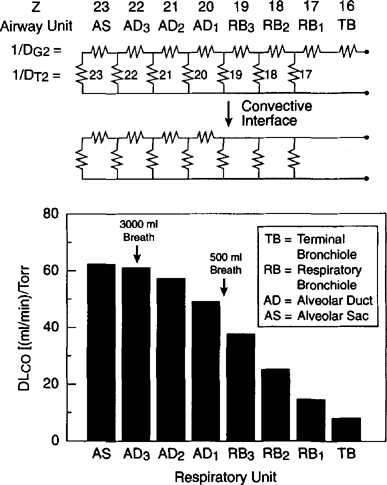
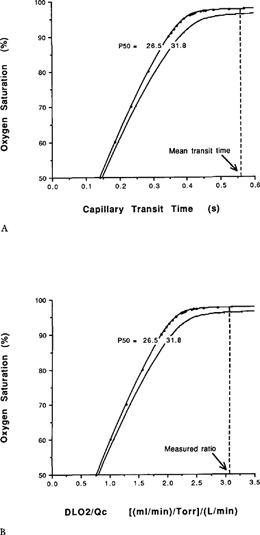
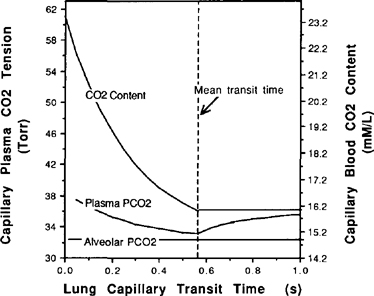
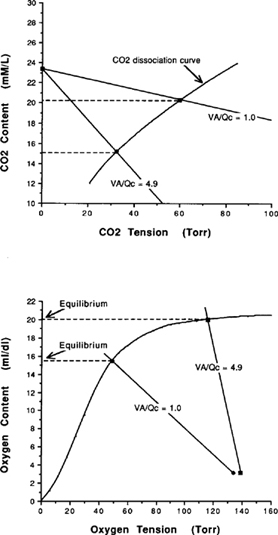
| References |
| 1. |
Adaro, F., and
L. E. Fahri.
Effects of intralobular gas diffustion on alveolar gas exchange.
Federation Proc.
30:
437,
1971 (Abstract).
|
| 2. |
Ahlborg, G.,
P. Felig,
L. Hagenfeldt,
R. Hendler, and
J. Wahren.
Substrate turnover during prolonged exercise in man. Splanchnic and leg metabolism of glucose, free fatty acids, and amino acids.
J. Clin. Invest.
53:
1080-1090,
1974.
|
| 3. |
Allen, C. J., and
N. L. Jones.
Rate of change in alveolar carbon dioxide and the control of ventilation during exercise.
J. Physiol. (Lond.)
355:
1-9,
1984.
|
| 4. |
Allen, T. A., and
W. S. Root.
Partition of carbon monoxide and oxygen between air and whole blood of rats, dogs and men as affected by plasma pH.
J. Appl. Physiol.
10:
186-190,
1957.
|
| 5. |
Altman, P. L., and
D. S. Dittman.
Respiration and Circulation, 2nd Ed. Biological Handbooks,
edited by P. L. Altman and
D. S. Dittmer.
Bethesda, MD:
Federation of American Societies for Experimental Biology,
1971,
p. 930.
|
| 6. |
Anderson, J. W., and
D. B. Jennings.
Osmolality, NaCl dietary intake, and regulation of ventilation by CO2.
Am. J. Physiol.
255
(Regulatory Integrative Comp. Physiol. 24):
R105-R112,
1988.
|
| 7. |
Åstrand, P.‐O.,
E. Hultman,
A. Juhlin‐Dannfelt, and
G. Reynolds.
Disposal of lactate during and after strenuous exercise in humans.
J. Appl. Physiol.
61:
338-343,
1986.
|
| 8. |
Astrup, P., and
J. W. Severinghaus.
History of Acid-Base Physiology.
Stockholm:
Munksgaard,
1986.
|
| 9. |
Bachofen, H.,
S. Schürch,
M. Urbinelli, and
E. R. Weibel.
Relations among alveolar surface tension, surface area, volume, and recoil pressure.
J. Appl. Physiol.
62:
1878-1887,
1987.
|
| 10. |
Bachofen, H.,
D. Wangensteen, and
E. R. Weibel.
Surfaces and volumes of alveolar tissue under zone II and zone III conditions.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
53:
879-885,
1982.
|
| 11. |
Bates, D. V.,
N. G. Boucot, and
A. E. Dormer.
The pulmonary diffusing capacity in normal subjects.
J. Physiol. (Lond.)
129:
237-252,
1955.
|
| 12. |
Bathe, K.
Finite Element Procedures in Engineering Analysis.
Englewood Cliffs, NJ:
Prentice‐Hall,
1932.
|
| 13. |
Bayly, W. M.,
B. D. Grant,
R. G. Breeze, and
J. W. Kramer.
The effects of maximal exercise on acid-base balance and arterial blood gas tension in thoroughbred horses.
In: Equine Exercise Physiology,
edited by D. H. Snow,
S. G. B. Persson, and
R. J. Rose.
Cambridge, UK:
Granta,
1983,
p. 400-407.
|
| 14. |
Bebout, D. E.,
D. Story,
J. Roca,
M. C. Hogan,
D. C. Poole,
C. R. Gonzalez,
O. Ueno,
P. Haab, and
P. D. Wagner.
Effects of altitude acclimatization on pulmonary gas exchange during exercise.
J. Appl. Physiol.
67:
2286-2295,
1989.
|
| 15. |
Bellingham, A. J.,
J. C. Detter, and
C. Lenfant.
Mechanisms of hemoglobin oxygen affinity in acidosis and alkalosis.
J. Clin. Invest.
50:
700-706,
1971.
|
| 16. |
Benade, A. J. S., and
N. Heisler.
Comparison of efflux rates of hydrogen and lactate ions from isolated muscles
in vitro. Respir. Physiol.
32:
369-380,
1978.
|
| 17. |
Bessman, S. P., and
F. Savabi.
The role of the phosphocreatine energy shuttle in exercise and muscular hypertrophy.
In: Biochemistry of Exercise VII,
edited by A. W. Taylor,
P. D. Gollnick,
H. J. Green,
C. D. Ianuzzo,
E. G. Noble,
G. Métivier, and
J. R. Sutton.
Champaign, IL:
Human Kinetics Press,
1990,
p. 167-178.
|
| 18. |
Bidani, A.,
E. D. Crandall, and
R. E. Forster.
Analysis of post capillary pH changes in blood in vivo after gas exchange.
J. Appl. Physiol.
44:
770-781,
1978.
|
| 19. |
Blanchard, E. M.,
B. Pan, and
R. J. Solero.
The effect of acidic pH on the ATPase activity and tronin CA2+ binding of rabbit skeletal myofilaments.
J. Biol. Chem.
259:
3181-3186,
1984.
|
| 20. |
Bohr, C.
Ueber die Lungenatmung.
Scand. Arch. Physiol.
2:
236-268,
1891.
|
| 21. |
Bohr, C.
Uber die spesifische Tatigkeit der Lungen bei der respiratorischen Gasaufnahme und ihr Verhalten zu der durch die alveolarwand stattfinden Gasdiffusion.
Scand. Arch. Physiol.
22:
221-280,
1909.
|
| 22. |
Bohr, C.,
K. A. Hasselbach, and
A. Krogh.
Uber inen in biologischen Beziehung wichtigen Einfluss, den die Kohlensaurespannung de Blutes auf dessen Saurstoffbindung ubt.
Scand. Arch. Physiol.
16:
401-412,
1904.
|
| 23. |
Borland, C. D. R., and
T. Higgenbottam.
A simultaneous single breath measurement of pulmonary diffusing capacity with nitric oxide and carbon monoxide.
Eur. Respir. J.
2:
56-63,
1989.
|
| 24. |
Boyden, E. A.
The structure of the pulmonary acinus in a child of six years and eight months.
Am. J. Anat.
132:
275-300,
1971.
|
| 25. |
Brooks, G. A.
The lactate shuttle during exercise and recovery.
Med. Sci. Sports Exerc.
18:
355-364,
1986.
|
| 26. |
Bunker, J. P.
The great trans‐Atlantic acid-base debate.
Anesthesia
26:
591-594,
1965.
|
| 27. |
Bur, S.,
H. Bachofen,
P. Gehr, and
E. R. Weibel.
Lung fixation by airway instillation: effects on capillary hematocrit.
Exp. Lung Res.
9:
57-66,
1985.
|
| 28. |
Burri, P. H., and
E. R. Weibel.
Morphometric estimation of pulmonary diffusion capacity. II. Effect of Po2 on the growing lung, adaption of the growing rat lung to hypoxia and hyperoxia.
Respir. Physiol.
11:
247-264,
1971.
|
| 29. |
Burt, C. T.,
J. Koutcher,
J. T. Roberts,
R. E. London, and
B. Chance.
Magnetic resonance spectroscopy of the musculoskeletal system.
Radiol. Clin. North Am.
24:
321-331,
1986.
|
| 30. |
Capen, R. L.,
L. P. Latham, and
W. W. Wagner, Jr..
Diffusing capacity of the lung during hypoxia: role of capillary recruitment.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
50:
165-171,
1981.
|
| 31. |
Carlin, J. I.,
C. C. W. Hsia,
S. S. Cassidy,
M. Ramanathan,
P. S. Clifford, and
R. L. Johnson, Jr..
Recruitment of lung diffusing capacity with exercise before and after pneumonectomy in dogs.
J. Appl. Physiol.
70:
135-142,
1991.
|
| 32. |
Cassoly, R., and
Q. H. Gibson.
Conformation, cooperativity and ligand binding in human hemoglobin.
J. Mol. Biol.
91:
301-313,
1975.
|
| 33. |
Chin, E. R.,
M. I. Lindinger, and
G. J. F. Heigenhauser.
Lactate metabolism in inactive skeletal muscle during lactacidosis.
Am. J. Physiol.
261
(Regulatory Integrative Comp. Physiol. 30):
R93-R105,
1991.
|
| 34. |
Chow, E. I.,
E. D. Crandall, and
R. E. Forster.
Kinetics of bicarbonate‐chloride exchange across the human red cell membrane.
J. Gen. Physiol.
68:
633-652,
1976.
|
| 35. |
Christianson, J.,
C. G. Douglas,
J. S. Haldane, and
J. B. S. Haldane.
The absorption and dissociation of carbon dioxide from human blood.
J. Physiol. (Lond.)
48:
244-271,
1914.
|
| 36. |
Christoforides, C.,
L. H. Laasberg, and
J. Hedley‐Whyte.
Effect of temperature on solubility of O2 in human plasma.
J. Appl. Physiol.
26:
56-60,
1969.
|
| 37. |
Clausen, T.
Significance of the Na+‐K+ pump regulation in skeletal muscle.
News Physiol. Sci.
5:
148-151,
1990.
|
| 38. |
Clode, M., and
E. J. M. Campbell.
The relationship between gas exchange and changes in blood lactate changes during exercise.
Clin. Sci.
37:
263-272,
1969.
|
| 39. |
Coates, G.,
H. O'Brodovich,
A. L. Jeffries, and
G. W. Gray.
Effects of exercise on lymph flow in sheep and goats during normoxia and hypoxia.
J. Clin. Invest.
74:
133-141,
1984.
|
| 40. |
Cohn, J. E.,
B. W. Carroll,
B. W. Armstrong,
R. H. Shephard, and
R. L. Riley.
Maximal diffusing capacity in the lung in normal male subjects of different ages.
J. Appl. Physiol.
6:
588-597,
1954.
|
| 41. |
Colebatch, H. J. H., and
C. K. Y. Ng.
Estimating alveolar surface area during life.
Respir. Physiol.
88:
163-170,
1992.
|
| 42. |
Constantine, H. P.,
M. R. Craw, and
R. E. Forster.
Rate of reaction of carbon dioxide with human red blood cells.
Am. J. Physiol.
208:
801-811,
1965.
|
| 43. |
Cooke, R.,
K. Franks,
G. B. Luciani, and
E. Pate.
The inhibition of rabbit skeletal muscle contraction by hydrogen ions and phosphate.
J. Physiol. (Lond.)
395:
77-97,
1988.
|
| 44. |
Crandall, E. D., and
J. E. Obrasky.
Direct evidence for participation of rat lung carbonic anhydrase in CO2 reactions.
J. Clin. Invest.
62:
618-622,
1978.
|
| 45. |
Crapo, J. D., and
R. O. Crapo.
Comparison of total lung diffusion capacity and the membrane component of diffusion capacity as determined by physiologic and morphometric techniques.
Respir. Physiol.
51:
183-194,
1983.
|
| 46. |
Crapo, J. D.,
R. O. Crapo,
R. L. Jensen,
R. R. Mercer, and
E. R. Weibel.
Evaluation of lung diffusing capacity by physiological and morphometric techniques [published erratum appears in J. Appl. Physiol. 65: following 2800, 1988.].
J. Appl. Physiol.
64:
2083-2091,
1988.
|
| 47. |
Craw, M. R.,
H. P. Constantine,
J. A. Morello, and
R. E. Forster.
Rate of the Bohr shift in human red cell suspensions.
J. Appl. Physiol.
18:
317-374,
1963.
|
| 48. |
Crawford, A. B. H.,
M. Makowska,
M. Paiva, and
L. A. Engel.
Convection and diffusion‐dependent ventilation maldistribution in normal subjects.
J. Appl. Physiol.
49:
838-846,
1985.
|
| 49. |
Cruz‐Orive, L. M., and
E. R. Weibel.
Recent stereological methods for cell biology: brief surgery.
Am. J. Physiol.
258
(Lung Cell. Mol. Physiol. 2):
L148-L156,
1990.
|
| 50. |
Cumming, G.,
J. Crank,
K. Horsefield, and
I. Parker.
Gaseous diffusion in the airways of the human lung.
Respir. Physiol.
1:
58-54,
1966.
|
| 51. |
Cumming, G.,
K. Horsfireld,
J. G. Jones, and
D. C. F. Muir.
The influence of gaseous diffusion on the alveolar plateau at different lung volumes.
Respir. Physiol.
2:
386-398,
1967.
|
| 52. |
Dalziel, K., and
J. C. Londesborough.
The mechanisms of reductive carboxylation reactions. Carbon dioxide or bicarbonate as substrate of nicotinamide‐adenine dinucleotide phosphate‐linked isocitrate dehydrogenase and malic enzyme.
Biochem. J.
110:
223-230,
1968.
|
| 53. |
Davies, G., and
L. Reid.
Growth of the alveoli and pulmonary arteries in childhood.
Thorax
25:
669-681,
1970.
|
| 54. |
Dawson, M. J.,
D. G. Gadian, and
D. R. Wilkie.
Muscular fatigue investigated by phosphorus nuclear magnetic resonance.
Nature
274:
861-865,
1978.
|
| 55. |
Dempsey, J. A.,
P. G. Hanson, and
K. S. Henderson.
Exercise induced arterial hypoxemia in healthy human subjects at sea level.
J. Physiol. (Lond.)
355:
161-175,
1984.
|
| 56. |
Douglas, A. R.,
N. L. Jones, and
J. W. Reed.
Calculation of whole blood CO2 content.
J. Appl. Physiol.
65:
473-477,
1988.
|
| 57. |
Dunnill, M. S.
Postnatal growth of the lung.
Thorax
17:
329-333,
1962.
|
| 58. |
Dupuis, J.,
C. A. Goresky,
C. Junear,
A. Calderone,
J. L. Rouleau,
C. P. Rose, and
S. Goresky.
Use of norepinephrine uptake to measure lung capillary recruitment with exercise.
J. Appl. Physiol.
68:
700-713,
1990.
|
| 59. |
Dupuis, J.,
C. A. Goresky,
J. W. Ryan,
J. L. Rouleau, and
G. G. Bach.
Pulmonary angiotensin‐converting enzyme substrate hydrolysis during exercise.
J. Appl. Physiol.
75:
1868-1886,
1992.
|
| 60. |
Edwards, R. H. T.
Human muscle function and fatigue.
Ciba Pound. Symp.
82:
1-18,
1981.
|
| 61. |
Effros, R. M.,
L. Shapiro, and
P. Silverman.
Carbonic anhydrase activity of rabbit lungs.
J. Appl. Physiol.
49:
589-600,
1980.
|
| 62. |
Eldridge, F. L.
Central integration of mechanisms in exercise dyspnea.
Med. Sci. Sports Exerc.
26:
319-327,
1994.
|
| 63. |
Evans, J. W., and
P. D. Wagner.
Limits on V.A/Q. distribution from analysis of experimental inert gas elimination.
J. Appl. Physiol.
42:
889-898,
1977.
|
| 64. |
Farrell, P. A.,
M. B. Maron,
L. H. Hamilton,
M. G. Maksud, and
C. Foster.
Time course of lung volume changes during prolonged treadmill exercise.
Med. Sci. Sports Exerc.
15:
319-324,
1983.
|
| 65. |
Federspiel, W. J.
Pulmonary diffusing capacity: implications of two‐phase blood flow in capillaries.
Respir. Physiol.
77:
119-134,
1989.
|
| 66. |
Figge, J.,
T. Mydosh, and
V. Fencl.
Serum proteins and acid-base equilibria: a follow‐up.
J. Lab. Clin. Med.
120:
713-719,
1992.
|
| 67. |
Filley, G. F.,
D. J. MacIntosh, and
G. W. Wright.
Carbon monoxide uptake and pulmonary diffusing capacity in normal subjects at rest and during exercise.
J. Clin. Invest.
33:
530-539,
1954.
|
| 68. |
Filley, G. F.,
G. D. Swanson, and
N. B. Kindig.
Chemical breathing controls: slow, intermediate and fast.
Clin. Chest Med.
1:
13-32,
1980.
|
| 69. |
Forrest, J. B., and
E. R. Weibel.
Morphometric estimation of pulmonary diffusion capacity. VII. The normal guinea pig lung.
Respir. Physiol.
24:
191-202,
1975.
|
| 70. |
Forster, R. E.
Exchange of gases between alveolar air and pulmonary capillary blood: pulmonary diffusing capacity.
Physiol. Rev.
37:
391-452,
1957.
|
| 71. |
Forster, R. E.
Diffusion of gases across the alveolar capillary membrane.
In: Handbook of Physiology, The Respiratory System, Gas Exchange,
edited by L. E. Fahri and
S. M. Tenney.
Washington, DC:
Am. Physiol. Soc.,
1987,
p. 71-88.
|
| 72. |
Fung, Y. C. B., and
S. S. Sobin.
Pulmonary alveolar blood flow.
In: Bioengineering Aspects of the Lung,
edited by J. B. West.
New York and Basel:
Marcel Dekker, Inc.,
1977,
p. 267-359.
|
| 73. |
Gale, G. E.,
J. R. Torre‐Bueno,
R. E. Moon,
H. A. Saltzman, and
P. D. Wagner.
Ventilation/perfusion inequality in normal humans during exercise at sea level and simulated altitude.
J. Appl. Physiol.
58:
978-988,
1985.
|
| 74. |
Gamble, J. L.
Chemical Anatomy, Physiology and Pathology of Extracellular Fluid.
Cambridge, MA:
Harvard University Press,
1949.
|
| 75. |
Geelhaar, A., and
E. R. Weibel.
Morphometric estimation of pulmonary diffusion capacity. 3. The effect of increased oxygen consumption in Japanese waltzing mice.
Respir. Physiol.
11:
354-366,
1971.
|
| 76. |
Gehr, P.,
M. Bachofen, and
E. R. Weibel.
The normal human lung: ultrastructure and morphometric estimation of diffusion capacity.
Respir. Physiol.
32:
121-140,
1978.
|
| 77. |
Gibson, Q. H.,
F. Kreuzer,
E. Medu, and
F. J. W. Roughton.
The kinetics of human hemoglobin in solution and in the red cell at 37°C.
J. Physiol. (Lond.)
129:
65-89,
1955.
|
| 78. |
Gibson, Q. H., and
F. J. W. Roughton.
The reactions of sheep hemoglobin with nitric oxide.
J. Physiol. (Lond.)
128:
69P,
1955.
|
| 79. |
Gil, J.,
H. Bachofen,
P. Gehr, and
E. R. Weibel.
Alveolar volume‐surface area relation in air‐ and saline‐filled lungs fixed by vascular perfusion.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
47:
990-1001,
1979.
|
| 80. |
Gladden, L. B.
Lactate uptake by skeletal muscle.
In: Exercise and Sports Sciences Reviews,
edited by K. B. Pandolf.
Baltimore:
Williams & Wilkins,
1989,
p. 115-116.
|
| 81. |
Gledhill, N.,
A. B. Froese, and
J. A. Dempsey.
Ventilation to perfusion distribution during exercise in health.
In: Muscular Exercise and the Lung,
edited by J. A. Dempsey and
C. E. Reed.
Madison, WI:
University of Wisconsin Press,
1977,
p. 325-344.
|
| 82. |
Graham, B. L.,
J. T. Mink, and
D. J. Cotton.
Improved accuracy and precision of single‐breath CO diffusing capacity measurements.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
51:
1306-1313,
1981.
|
| 83. |
Gros, G., and
S. J. Dodgson.
Velocity of CO2 exchange in muscle and liver.
Ann. Rev. Physiol.
50:
669-694,
1988.
|
| 84. |
Grote, J.
Die Sauerstoffeddiffusionscostanten im lungengewebe and wasser und ihre temperaturebhangikeit.
Pflugers Arch. Ges. Physiol.
295:
245-254,
1967.
|
| 85. |
Guenard, H.,
N. Varene, and
P. Vaida.
Determination of lung capillary blood volume and membrane diffusing capacity in patients in man by the measurements of NO and CO transfer.
Respir. Physiol.
70:
113-120,
1987.
|
| 86. |
Gundersen, H. J. G.,
T. B. Jensen, and
R. Østerby.
Distribution of membrane thickness determined by lineal analysis.
J. Microsc.
113:
27-43,
1978.
|
| 87. |
Gutierrez, G., and
J. J. Ronco.
Tissue gas exchange.
In: Pulmonary and Critical Care Medicine,
edited by R. C. Bone.
St Louis:
Mosby,
1994,
p. 1-18.
|
| 88. |
Haefeli, B. B., and
E. R. Weibel.
Morphometry of the human pulmonary acinus.
Anat. Rec.
220:
401-414,
1988.
|
| 89. |
Haldane, J., and
J. L. Smith.
The oxygen tension of arterial blood.
J. Physiol. (Lond.)
20:
497-518,
1896.
|
| 90. |
Hammond, M. D.,
G. E. Gale,
K. S. Kapitan,
A. Ries, and
P. D. Wagner.
Pulmonary gas exchange in humans during normobaric hypoxic exercise.
J. Appl. Physiol.
61:
1749-1757,
1986.
|
| 91. |
Hammond, M. D., and
S. C. Hempleman.
Oxygen diffusing capacity estimates derived from measured VA/Q distributions in man.
Respir. Physiol.
69:
129-147,
1987.
|
| 92. |
Hansen, J. E., and
E. P. Ampaya.
Human air space shapes, sizes, areas and volumes.
J. Appl. Physiol.
38:
990-995,
1975.
|
| 93. |
Hartridge, H., and
F. J. W. Roughton.
A method for measuring the velocity of very rapid reactions.
Proc. R. Soc. Lond. A
104:
376-394,
1923.
|
| 94. |
Hartridge, H., and
F. J. W. Roughton.
The velocity with which carbon monoxide displaces oxygen from combination with hemoglobin. Part I.
Proc. R. Soc. Lond., B
94:
336-367,
1923.
|
| 95. |
Hartridge, H., and
F. J. W. Roughton.
The velocity with which oxygen combines with reduced hemoglobin. Part III.
Proc. R. Soc. Lond. A
107:
654-683,
1925.
|
| 96. |
Hasselbalch, K.
Neutralitatsregulation und reizbarkeit des atemzentrums in ihren Wirkungen auf die koklensaurespannung des Blutes.
Bioochem. Z.
46:
403-439,
1912.
|
| 97. |
Havel, R. J.,
B. Pernow, and
N. L. Jones.
Uptake and release of fatty acids and other metabolites in legs of exercising men.
J. Appl. Physiol.
23:
90-99,
1967.
|
| 98. |
Heidelberger, E., and
R. B. Reeves.
O2 transfer kinetics in a whole blood unicellular thin layer.
J. Appl. Physiol.
68:
1854-1890,
1990.
|
| 99. |
Heigenhauser, G. J. F.,
N. L. Jones,
J. M. Kowalchuk, and
M. I. Lindinger.
The role of the physicochemical systems in plasma in acid-base control in exercise.
In: Biochemistry of Exercise VII,
edited by A. W. Taylor,
P. D. Gollnick,
H. J. Green,
C. D. Ianuzzo,
E. G. Noble,
G. Metivier, and
J. R. Sutton.
Champaign, IL:
Human Kinetics,
1990,
p. 359-374.
|
| 100. |
Heigenhauser, G. J. F., and
M. I. Lindinger.
The total ionic status of muscle during intense exercise.
In: Oxygen Transfer from Atmosphere to Tissues,
edited by N. C. Gonzales and
M. R. Fedde.
New York:
Plenum Press,
1988,
p. 237-242.
|
| 101. |
Heisler, N., and
J. Piiper.
The buffer value of rat diaphragm muscle tissue determined by Pco2 equilibration of hemogenates.
Respir. Physiol.
12:
169-178,
1971.
|
| 102. |
Heisler, N., and
J. Piiper.
Determination of intracellular buffering properties in rat diaphragm muscle.
Am. J. Physiol.
222:
747-753,
1972.
|
| 103. |
Hemingway, A.,
C. J. Hemingway, and
F. J. W. Roughton.
The rate of chloride shift of respiration studied with a rapid filtration method.
Respir. Physiol.
10:
1-9,
1970.
|
| 104. |
Hempleman, S. C., and
A. T. Gray.
Estimating steady‐state DLO2 with nonlinear dissociation curves and Va/Q inequality.
Respir. Physiol.
732:
279-288,
1988.
|
| 105. |
Henderson, L. J.
Blood: A Study in General Physiology.
New Haven, CT:
Yale University Press,
1928.
|
| 106. |
Hermansen, L., and
J.‐B. Osnes.
Blood and muscle pH after maximal exercise in man.
J. Appl. Physiol.
32:
304-308,
1972.
|
| 107. |
Hermansen, L., and
Stensvold.
Production and removal of lactate during exercise in man.
Acta Physiol. Scand.
86:
191-201,
1972.
|
| 108. |
Hermansen, L., and
O. Vaage.
Lactate disappearance and glycogen synthesis in human muscle after maximal exercise.
Am. J. Physiol.
233
(Endocrinol. Metab. Gastrointest. Physiol. 2):
E422-E429,
1977.
|
| 109. |
Hill, A. V.
The possible effects of the aggregation of the hemoglobin molecules of hemoglobin on the dissociation curve.
J. Physiol. (Lond.)
40:
iv-vii,
1910.
|
| 110. |
Hill, E. P.,
G. G. Power, and
R. D. Gilbert.
Mathematical simulation of pulmonary O2 and CO2 exchange.
Am. J. Physiol.
224:
904-917,
1973.
|
| 111. |
Hill, E. P.,
G. G. Power, and
R. D. Gilbert.
Rate of pH changes in blood plasma in vitro and in vivo.
J. Appl. Physiol
42:
928-934,
1977.
|
| 112. |
Hill, R. J. W.,
G. Konigsberg,
G. Guidotti, and
L. C. Craig.
The structure of human hemoglobin I. The separation of the α and β‐ chains and their amino acid composition.
J. Biol. Chem.
237:
1549-1554,
1962.
|
| 113. |
Hlastala, M. P.,
P. Scheid, and
J. Piiper.
Interpretation of inert gas retention and excretion in the presence of stratified inhomogeneity.
Respir. Physiol.
46:
247-259,
1981.
|
| 114. |
Hochachka, P. W., and
G. N. Somero.
Biochemical Adaptation.
Princeton, NJ:
Princeton University Press,
1984.
|
| 115. |
Hogan, M. C.,
D. E. Bebout,
A. T. Gray,
P. D. Wagner,
J. B. West, and
P. E. Haab.
Muscle maximal O2 uptake at constant O2 delivery with and without CO in the blood.
J. Appl. Physiol.
69:
830-836,
1990.
|
| 116. |
Holland, R. A.,
H. Shibata,
P. Scheid, and
J. Piiper.
Kinetics of O2 uptake and release by red cells in stopped‐flow apparatus: effects of unstirred layer.
Respir. Physiol.
59:
71-91,
1985.
|
| 117. |
Holland, R. A. B.
Rate at which CO replaces O2 from O2 Hb in red cells of different species.
Respir. Physiol.
7:
43-63,
1969.
|
| 118. |
Holland, R. A. B.,
W. Van Hezewijk, and
J. Zubzanda.
Velocity of oxygen uptake by partly saturated adult and fetal human red cells.
Respir. Physiol.
29:
303-314,
1977.
|
| 119. |
Honig, C. L.,
T. E. J. Gayeski,
W. J. Federspiel,
A. Clark, Jr., and
P. Clark.
Gradients from hemoglobin to cytochrome: new concepts, new complexities.
Adv. Exp. Med. Biol.
169:
23-38,
1984.
|
| 120. |
Hopkins, S. R.,
A. S. Belzberg,
B. R. Wiggs, and
D. C. McKenzie.
Pulmonary transit time and diffusion limitation during heavy exercise in athletes.
Respir. Physiol.
1995
(in press).
|
| 121. |
Hsia, C. C. W.,
C. J. C. Chuong, and
R. L. Johnson, Jr..
Critique of the conceptual basis of diffusing capacity estimates: a finite element analysis.
J. Appl. Physiol.
1995
(in press).
|
| 122. |
Hsia, C. C. W.,
F. Fryder‐Doffey,
V. Stalder‐Navarro,
J. Johnson,
R. C. Reynods, and
E. R. Weibel.
Structural changes underlying compensatory increase of diffusing capacity after left pneumonectomy in adult dogs.
J. Clin. Invest.
92:
758-764,
1993.
|
| 123. |
Hsia, C. C. W.,
L. F. Herazo,
F. Frydor‐Doffey, and
E. R. Weibel.
Compensatory lung growth occurs in dogs after right pneumonectomy.
J. Clin. Invest.
94:
758-764,
1994.
|
| 124. |
Hsia, C. C. W.,
L. F. Herazo,
M. Ramanathan, and
R. L. Johnson, Jr..
Cardiopulmonary adaptations to pneumonectomy in dogs. IV Total diffusing capacity, membrane diffusing capacity and capillary blood volume.
J. Appl. Physiol.
77:
998-1005,
1994.
|
| 125. |
Hsia, C. C. W.,
L. F. Herazo,
M. Ramanathan, and
R. L. Johnson, Jr..
Cadiac output during exercise measured by acetylene rebreathing, thermodilution and Fick techniques.
J Appl. Physiol.
78:
1612-1616,
1995.
|
| 126. |
Hsia, C. C. W.,
L. F. Herazo,
M. Ramanathan,
R. L. Johnson, Jr., and
P. D. Wagner.
Cardiopulmonary adaptations to pneumonectomy in dogs. II. Ventilation‐perfuson relationships and microvascular recruitment.
J. Appl. Physiol
74:
1299-1309,
1993.
|
| 127. |
Hsia, C. C. W.,
M. Ramanathan, and
A. S. Estrera.
Recruitment of diffusing capacity with exercise in patients after pneumonectomy.
Am. Rev. Respir. Dis.
145:
811-816,
1992.
|
| 128. |
Huang, Y. T.,
M. J. Helms, and
N. R. MacIntyre.
Normal values for single exhalation diffusing capacity and pulmonary capillary blood flow in sitting, supine positions and during mild exercise.
Chest
105:
501-508,
1994.
|
| 129. |
Hufner, G.
Neue Versuche zu Bestimmung der Saurstoffecapacitat des Blutfarbstoffes.
Arch. Physiol. (Leipzig):
130-176,
1894.
|
| 130. |
Hultman, E., and
K. Sahlin.
Acid-base balance during exercise.
Exerc. Sport Sci. Rev.
8:
41-127,
1980.
|
| 131. |
Jacobs, M. H.
The production of intracellular acidity by neutral and alkaline solutions containing carbon dioxide.
Am. J. Physiol.
53:
457-463,
1920.
|
| 132. |
Johnson, R. L., Jr., and
C. C. W. Hsia.
Functional recruitment of pulmonary capillaries.
J. Appl. Physiol.
76:
1405-1407,
1994.
|
| 133. |
Johnson, R. L., Jr., and
M. Ramanathan.
Buffer equilibria in the lungs.
In: the Kidney, Physiology and Pathophysiology,
edited by D. W. Seldin and
G. Giebisch.
New York:
Raven Press,
1985,
p. 193-218.
|
| 134. |
Jones, N. L.,
G. J. F. Heigenhauser,
A. Kuksis,
C. G. Matsos,
J. R. Sutton, and
C. J. Toews.
Fat metabolism in heavy exercise.
Clin. Sci.
59:
469-478,
1980.
|
| 135. |
Jones, N. L.,
N. McCartney,
T. Graham,
L. L. Spriet,
J. M. Kowalchuk,
G. J. F. Heigenhauser, and
J. R. Sutton.
Muscle performance and metabolism in maximal isokinetic cycling at slow and fast speeds.
J. Appl. Physiol.
59:
132-136,
1985.
|
| 136. |
Jones, R. S., and
F. A. Meade.
A theoretical and experimental analysis of anomalies in the estimation of pulmonary diffusing capacity by the single breath method. Q.
J. Exp. Physiol.
46:
131-143,
1961.
|
| 137. |
Juel, C.
Potassium and sodium shifts during in vitro isometric muscle contraction, and the time course of the ion‐gradient recovery.
Pflugers Arch.
406:
458-463,
1986.
|
| 138. |
Juel, C.
Intracellular pH recovery and lactate efflux in mouse soleus muscles stimulated in vitro: the involvement of sodium/proton exchange and a lactate carrier.
Acta Physiol. Scand.
132:
363-371,
1988.
|
| 139. |
Karas, R. H.,
C. R. Taylor,
J. H. Jones,
S. L. Lindstedt,
R. B. Reeves, and
E. R. Weibel.
Adaptive variation in the mammalian respiratory system in relation to energetic demand: VII. Flow of oxygen across the pulmonary gas exchanger.
Respir. Physiol.
69:
101-115,
1987.
|
| 140. |
Kelman, G. R.
Digital computer procedure for the conversion of PCO2 into blood content.
Respir. Physiol.
3:
111-115,
1967.
|
| 141. |
Kety, S.
The theory and applications of the exchange of inert gas at the lungs and tissues.
Pharmacol. Rev.
3:
1-41,
1951.
|
| 142. |
Klocke, R. A.
Mechanism and kinetics of the Haldane effect in human erythrocytes.
J. Appl. Physiol.
35:
673-681,
1973.
|
| 143. |
Klocke, R. A.
Mechanism and kinetics of the Haldane effect on human erythrocytes.
J. Appl. Physiol.
35:
678-681,
1973.
|
| 144. |
Klocke, R. A.
Rate of bicarbonate‐chloride exchange in human red cells at 37°C.
J. Appl. Physiol.
40:
707-714,
1976.
|
| 145. |
Klocke, R. A.
Catalysis of CO2 reactions by lung carbonic anhydrase.
J. Appl. Physiol.
44:
882-888,
1978.
|
| 146. |
Klocke, R. A.
Velocity of CO2 exchange in blood.
Ann. Rev. Physiol.
50:
625-637,
1988.
|
| 147. |
Konig, M. F.,
J. M. Lucocq, and
E. R. Weibel.
Demonstration of pulmonary vascular perfusion by electron and light microscopy.
J. Appl. Physiol.
75:
1877-1883,
1993.
|
| 148. |
Kowalchuk, J. M.,
G. J. F. Heigenhauser,
M. I. Lindinger,
G. Obminski,
J. R. Sutton, and
N. L. Jones.
Role of lungs and inactive muscle in acid-base control after maximal exercise.
J. Appl. Physiol.
65:
2090-2096,
1988.
|
| 149. |
Kowalchuk, J. M.,
G. J. F. Heigenhauser,
M. I. Lindinger,
J. R. Sutton, and
N. L. Jones.
Factors influencing hydrogen ion concentration in muscle after intense exercise.
J. Appl. Physiol.
65:
2080-2089,
1988.
|
| 150. |
Kowalchuk, J. M.,
G. J. F. Heigenhauser,
J. R. Sutton, and
N. L. Jones.
Effect of chronic acetazolamide administration on gas exchange and acid-base control after maximal exercise.
J. Appl Physiol.
76:
1211-1219,
1994.
|
| 151. |
Kowalchuk, J. M.,
G. J. F. Heigenhauser,
J. R. Sutton, and
N. L. Jones.
Effect of acetazolamide on gas exchange and acid-base control after maximal exercise.
J. Appl. Physiol.
72:
278-287,
1992.
|
| 152. |
Kowalchuk, J. M., and
B.W. Scheuermann.
Acid-base regulation: a comparison of quantitative methods.
Can. J. Physiol. Pharmacol.
72:
818-826,
1994.
|
| 153. |
Krebs, H. A., and
F. J. W. Roughton.
Carbonic anhydrase as a tool in studying the mechanisms of reactions involving H2 CO3, CO2 or HCO3−
Biochem. J.
43:
550-555,
1948.
|
| 154. |
Krogh, A., and
M. Krogh.
On the rate of diffusion of carbonic oxide into the lungs of man.
Scand. Arch. Physiol.
23:
236-247,
1909.
|
| 155. |
Langston, C.,
K. Kida,
M. Reed, and
W. M. Thurlbeck.
Human lung growth in late gestation and in the neonate.
Am. Rev. Respir. Dis.
129:
607-613,
1984.
|
| 156. |
Larsen, L. A., and
J. M. Burnell.
Muscle buffer values.
Am. J. Physiol.
234
(Renal Fluid Electrolyte Physiol. 3):
F432-F436,
1978.
|
| 157. |
Lewis, B. M.,
T. Lin,
F. E. Noe, and
E. J. Hayford‐Welsing.
the measurement of pulmonary diffusing capacity for carbon monoxide by a rebreathing method.
J. Clin. Invest.
38:
2073-2086,
1959.
|
| 158. |
Lilienthal, J. L., Jr.,
R. L. Riley,
D. D. Proemmel, and
R. E. Franke.
An experimental analysis in man of the pressure gradient from alveolar air to arterial blood during rest and exercise at sea level and at high altitude.
Am. J. Physiol.
147:
199-216,
1946.
|
| 159. |
Lindinger, M. I., and
G. J. F. Heigenhauser.
Acid-base systems in skeletal muscle and their response to exercise.
In: Biochemistry of Exercise VII,
edited by P. D. Gollnick,
A. W. Taylor,
H. J. Green,
C. D. Ianuzzo,
E. G. Noble,
G. Metivier, and
J. R. Sutton.
Champaign, IL:
Human Kinetics,
1990,
p. 341-357.
|
| 160. |
Lindinger, M. I.,
G. J. F. Heigenhauser,
R. S. McKelvie, and
N. L. Jones.
Blood ion regulation during repeated maximal exercise and recovery in humans.
Am. J. Physiol.
262
(Regulatory Integrative Comp. Physiol. 31):
R126-R136,
1992.
|
| 161. |
Lindinger, M. I.,
G. J. F. Heigenhauser, and
L. L. Spriet.
Effects of alkalosis on muscle ions at rest and with intense exercise.
Can. J. Physiol. Pharmacol.
68:
820-829,
1990.
|
| 162. |
Lindinger, M. I.,
L. L. Spriet,
E. Hultman,
T. Putman,
R. S. McKelvie,
L. C. Lands,
N. L. Jones, and
G. J. F. Heigenhauser.
Plasma volume and ion regulation during exercise after low‐ and high‐carbohydrate diets.
Am. J. Physiol.
266
(Regulatory Integrative Comp. Physiol. 35):
R1896-R1906, 1994.
|
| 163. |
MacIntyre, N. R., and
J. A. Nadel.
Regional diffusing capacity in normal lungs during a slow exhalation.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
52:
1487-1492,
1983.
|
| 164. |
Manier, G.,
J. Moinard,
P. Techoueryres,
N. Varene, and
H. Guenard.
Pulmonary diffusion limitation after prolonged strenuous exercise.
Respir. Physiol.
83:
143-154,
1991.
|
| 165. |
Maron, M. B., and
L. H. Hamilton, and
M. G. Maksud.
Alterations in pulmonary function consequent to competitive marathon running.
Med. Sci. Sports Exerc.
11:
244-249,
1979.
|
| 166. |
Martonen, T. B., and
A. F. Wilson.
Theoretical basis of single breath gas absorption tests.
J. Math. Biol.
14:
203-220,
1982.
|
| 167. |
McKelvie, R. S.,
N. L. Jones, and
G. J. F. Heigenhauser.
Factors contributing to increased muscle fatigue with β‐blockers.
Can. J. Physiol. Pharmacol.,
69:
254-261,
1991.
|
| 168. |
McKelvie, R. S.,
M. I. Lindinger,
G. J. F. Heigenhauser, and
N. L. Jones.
Contribution of erythrocytes to the control of the electrolyte changes of exercise.
Can. J. Physiol. Pharmacol.
69:
984-993,
1996.
|
| 169. |
Medbd, J. I., and
O. M. Sejersted.
Acid-base and electrolyte balance after exhausting exercise in endurance‐trained and sprint‐trained subjects.
Acta Physiol. Scand.
125:
97-109,
1985.
|
| 170. |
Medbo, J. I., and
O. M. Sejersted.
Plasma potassium changes with high intensity exercise.
J. Physiol. (Lond.)
421:
105-122,
1990.
|
| 171. |
Meldrum, N. U., and
F. J. W. Roughton.
Carbonic anhydrase. Its preparation and properties.
J. Physiol. (Lond.)
80:
113-142,
1933.
|
| 172. |
Merlot‐Benichou, C.,
M. Sinet,
M. C. Blayo, and
C. Gaudebout.
Oxygen‐combining capacity in dog. In vitro and in vivo determination.
Respir. Physiol.
21:
87-99,
1974.
|
| 173. |
Meyer, M.,
K. Schuster,
H. Schulz,
M. Mohr, and
J. Piiper.
Pulmonary diffusing capacities for nitric oxide and carbon monoxide determined by rebreathing in dogs.
J. Appl. Physiol.
68:
2344-2357,
1990.
|
| 174. |
Miles, D. S., and
R. J. Durbin.
Alterations in pulmonary function consequent to a 5‐mile run.
J. Sports Med.
25:
90-97,
1985.
|
| 175. |
Miller, J. M., and
R. L. Johnson, Jr..
Effect of lung inflation on pulmonary diffusing capacity at rest and exercise.
J. Clin. Invest.
45:
493-500,
1966.
|
| 176. |
Monod, J.,
J. Wyman, and
J. Changeux.
One the nature of allosteric transitions: a plausible model.
J. Mol. Biol.
12:
88-118,
1965.
|
| 177. |
Nattie, E. E., and
Y. N. Cai.
CSF and plasma ions and blood gases during organic metabolic acidosis in conscious rabbits.
J. Appl. Physiol.
57:
68-76,
1984.
|
| 178. |
Needham, D.
Machina Carnis. The Biochemistry of Muscular Contraction in its Historical Development.
New York:
Cambridge University Press,
1976.
|
| 179. |
Newth, C. J. L.,
D. J. Cotton, and
J. A. Nadel.
Pulmonary diffusing capacity measured at multiple intervals during a slow exhalation in man.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
43:
617-625,
1977.
|
| 180. |
Nicholson, P., and
F. J. W. Roughton.
A theoretical study of the influence of diffusion and chemical reaction velocity on the rate of exchange of carbon monoxide and oxygen between the red blood corpuscle and the surrounding fluid.
Proc. R. Soc. Lond. B
138:
241-264,
1951.
|
| 181. |
Ogilvie, C. M.,
R. E. Forster,
W. S. Blakemore, and
J. W. Morton.
A standardized breath holding technique for the clinical measurement of the diffusing capacity of the lung for carbon monoxide.
J. Clin. Invest.
36:
1-17,
1957.
|
| 182. |
Otis, A. B.
Quantitative relationships in steady‐state gas exchange.
In: Handbook of Physiology, Respiration,
edited by W. O. Fenn and
H. Rahn,
Washington, DC:
Am. Physiol. Soc.,
1964,
p. 681-698.
|
| 183. |
Paiva, M.
Gas transport in the lung.
J. Appl. Physiol.
35:
401-410,
1973.
|
| 184. |
Paiva, M.
Gas mixing and distribution in the human lung.
In: Lung Biology in Health and Disease,
edited by L. A. Engel and
M. Paiva.
New York:
Marcel Dekker, Inc.,
1985,
p. 221-285.
|
| 185. |
Paiva, M.,
S. Verbank, and
A. van Muylem.
Diffusion‐dependent contribution to the slope of the alveolar plateau.
Respir. Physiol.
72:
257-270,
1988.
|
| 186. |
Pannier, J. L.,
J. Weyne, and
I. Leusen.
Effects of PCO2, bicarbonate and lactate on the isometric contractions of isolated soleus muscle of the rat.
Pflugers Arch.
320:
120-132,
1970.
|
| 187. |
Park, H. K., and
R. Reeves.
Thin film measurements of Q(CO) in human red cells at zero, partial and full oxygen saturation.
FASEB J.
4:
901,
1990.
|
| 188. |
Parker, H.,
K. Horsefield, and
G. Cumming.
Morphology of distal airways in the human lung.
J. Appl. Physiol.
31:
386-391,
1971.
|
| 189. |
Patterson, D. J.
Potassium and ventilation in exercise.
J. Appl. Physiol.
72:
811-820,
1992.
|
| 190. |
Persson, S. G. B.,
P. Railings, and
C. Ingvast‐Larsson.
Relationsips between arterial oxygen tension and cardiopulmonary function during submaximal exercise in the horse.
In: Equine Exercise Physiology 2,
edited by J. R. Gillespie,
N. E. Robinson, and
C. A. Davis.
Davis, CA:
ICEEP Publications,
1987,
p. 161-171.
|
| 191. |
Perutz, M. F.
Stereochemistry of cooperative effects in hemoglobin.
Nature
228:
726-739,
1970.
|
| 192. |
Pieschl, R. L.,
P. W. Toll,
D. E. Leith,
L. J. Peterson, and
M. R. Fedde.
Acid-base changes in the running greyhound: contributing variables.
J. Appl. Physiol.
73:
2297-2304,
1992.
|
| 193. |
Piiper, J., and
P. Scheid.
Model for capillary‐alveolar equilibration with special refernce to O2 uptake in hypoxia.
Respir. Physiol.
46:
193-208,
1981.
|
| 194. |
Piiper, J., and
P. Scheid.
Models for a comparative functional analysis of gas exchange organs in vertebrates.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
53:
1321-1329,
1982.
|
| 195. |
Pitts, R. F.
The role of ammonia production and excretion in regulation of acid-base balance.
N. Engl. J. Med.
284:
32-38,
1971.
|
| 196. |
Podolsky, A.,
R. S. Eldridge,
R. S. Richardson,
D. R. Knight,
E. C. Johnson,
S. R. Hopkins,
B. Michimata,
B. Grassi,
S. S. Feiner,
S. S. Kurdak,
J. M. Uribe,
D. C. Poole,
P. E. Bickler,
J. W. Severinghause, and
P. D. Wagner.
Relationship between susceptibility to high altitude pulmonary edema and to exercise induced ventilation/perfusion mismatch.
Am. J. Respir. Crit. Care Med.
149:
A818,
1994.
|
| 197. |
Powers, G. P.
Solubility of O2 and CO in blood and pulmonary and placental tissue.
J. Appl. Physiol.
24:
468-474,
1968.
|
| 198. |
Presson, R. G.,
C. C. Hanger,
P. S. Godbey,
J. A. Graham,
T. C. Lloyd, Jr., and
W. W. Wagner, Jr..
Effect of increasing flow on distribution of pulmonary transit times.
J. Appl. Physiol.
76:
1701-1711,
1994.
|
| 199. |
Pump, K. K.
Morphology of the acinus of the human lung.
Dis. Chest
56:
126-134,
1969.
|
| 200. |
Putman, C. T.,
L. L. Spriet,
E. Hultman,
M. I. Lindinger,
L. C. Lands,
R. S. McKelvie,
G. Cederblad,
N. L. Jones, and
G. J. F. Heigenhauser.
Pyruvate dehydrogenase activity and acetyl group accumulation during exercise after different diets.
Am. J. Physiol.
265
(Endocrinol. Metab. 28):
E752-E760,
1993.
|
| 201. |
Radda, G. K.
The use of NMR spectroscopy for the understanding of disease.
Science
233:
640-645,
1986.
|
| 202. |
Rahn, H., and
W. O. Fenn.
A Graphical Analysis of Respiratory Gas Exchange: The O2-CO2 Diagram.
Washington, DC:
Am. Physiol. Soc.,
1955.
|
| 203. |
Rahn, H.,
R. B. Reeves, and
B. J. Howell.
Hydrogen ion regulation, temperature and evolution.
Am. Rev. Respir. Dis.
112:
165-172,
1975.
|
| 204. |
Ratner, E. R., and
P. D. Wagner.
Resolution of the multiple inert gas method for estimating VA/Q distributions.
Respir. Physiol.
49:
293-313,
1982.
|
| 205. |
Rauwerda, P. E.
Unequal Ventilation of Different Parts of the Lung and Determination of Cardiac Output
(thesis). University of Groningen,
1946.
|
| 206. |
Reeves, R. B.
An imidazole alphastat hypothesis for vertebrate acid-base regulation: tissue carbon dioxide content and body temperature in bullfrogs.
Respir. Physiol.
14:
219-236,
1972.
|
| 207. |
Reeves, R. B., and
H. K. Park.
CO uptake kinetics of red cells and CO diffusing capacity.
Respir. Physiol.
88:
1-21,
1992.
|
| 208. |
Riley, R. L., and
A. Cournand.
“Ideal” alveolar air and analysis of ventilation‐perfusion relationships in the lungs.
J. Appl. Physiol.
1:
825-847,
1949.
|
| 209. |
Robertson, H. T.,
J. Whitehead, and
M. P. Hlastala.
Diffusion‐related differences in elimination of inert gases from the lung.
J. Appl. Physiol.
61:
1162-1172,
1986.
|
| 210. |
Rose, R. J.,
D. R. Hodgson,
T. B. Kelso,
L. J. McCutcheon,
W. M. Bayly, and
P. D. Gollnick.
Effects of acetazolamide on metabolic and respiratory responses to exercise at maximal O2 uptake.
J. Appl. Physiol.
68:
617-626,
1990.
|
| 211. |
Rossing, T. H.,
N. Maffeo, and
V. Fencl.
Acid-base effects of altering plasma protein concentration in human blood in vitro.
J. Appl. Physiol.
61:
2260-2265,
1986.
|
| 212. |
Roughton, F. J. W.,
R. C. Darling, and
W. S. Root.
Factors affecting the determination of oxygen capacity, content and pressure in human arterial blood.
Am. J. Physiol.
147:
708-720,
1944.
|
| 213. |
Roughton, F. J. W., and
R. E. Forster.
Relative importance of diffusion and chemical reaction rates in determining the rate of exchange of gases in the human lung, with special reference to true diffusing capacity of the pulmonary membrane and volume of bood in lung capillaries.
J. Appl. Physiol.
11:
290-302,
1957.
|
| 214. |
Roughton, F. J. W.,
R. E. Forster, and
L. Cander.
Rate at which carbon monoxide replaces oxygen from combination with human hemoglobin in solution and in the red cell.
J. Appl. Physiol.
11:
269-276,
1957.
|
| 215. |
Rowell, L. B.,
H. L. Taylor,
Y. Wang, and
W. S. Carlson.
Saturation of arterial blood with oxygen during maximal exercise.
J. Appl. Physiol.
19:
284-286,
1964.
|
| 216. |
Sackner, M. A.,
D. Greenletch,
M. Heiman,
S. Epstein, and
N. Atkins.
Diffusing capacity, membrane diffusing capacity, capillary blood volume, pulmonary tissue volume and cardiac output by a rebreathing technique.
Am. Rev. Respir. Dis.
111:
157-165,
1975.
|
| 217. |
Saltin, B.,
R.F. Grover,
C. G. Blomqvist,
H. Hartley, and
R. L. Johnson, Jr..
Maximal oxygen uptake and cardiac output after 2 weeks at 4,300 m.
J. Appl. Physiol.
25:
400-409,
1968.
|
| 218. |
Schaffartzik, W.,
D. C. Poole,
T. Derion,
K. Tsukimoto,
M. C. Hogan,
J. P. Arcos,
D. E. Bebout, and
P. D. Wagner.
Va/Q distribution during heavy exercise and recovery in humans: implications for pulmonary edema.
J. Appl. Physiol.
72:
1657-1667,
1992.
|
| 219. |
Schaffartzik, W. J.,
J. Arcos,
K. Tsukimoto,
O. Mathieu‐Costello, and
P. D. Wagner.
Pulmonary interstitial edema in the pig after heavy exercise.
J. Appl. Physiol.
74:
2586-2594,
1993.
|
| 220. |
Scheid, P.,
M. P. Hlastala, and
J. Piiper.
Inert gas elimination from lungs with stratified in homogeneity: theory.
Respir. Physiol.
44:
299-309,
1981.
|
| 221. |
Scherer, P. W.,
S. Gobran,
S. J. Aukburg,
J. E. Baumgardner,
R. Bartkowski, and
G. R. Neufeld.
Numerical and experimental study of steady‐state CO2 and the inert gas washout.
J. Appl. Physiol.
64:
1022-1029,
1988.
|
| 222. |
Scherer, P. W.,
L. H. Schendalman, and
N. M. Greene.
Simultaneous diffusion and convection in single breath lung washout.
Bull. Math. Biophys.
34:
393-412,
1972.
|
| 223. |
Scholander, P. F.
Oxygen transport through hemoglobin solutions.
Science
131:
585-590,
1960.
|
| 224. |
Schreider, J. P., and
O. G. Raabe.
Structure of the human respiratory acinus.
Am. J. Nat.
162:
221-232,
1981.
|
| 225. |
Schwartz, W. B., and
A. S. Relman.
A critique of the parameters used in the evaluation of acid-base disorders. “Whole‐blood buffer base” and “standard bicarbonate” compared with blood pH and plasma bicarbonate concentration.
N. Engl. J. Med.
268:
1382-1388,
1963.
|
| 226. |
Sendroy, J., Jr.,
R. T. Dillon, and
D. D. Van Slyke.
Studies of gas and electrolyte equilibria in blood. XIX. The solubility and physical state of uncombined oxygen in blood.
J. Biol. Chem.
105:
597-632,
1934.
|
| 227. |
Severinghaus, J. W.
Blood gas calculator.
J. Appl. Physiol.
21:
1108-1116,
1966.
|
| 228. |
Shepard, R. H.,
E. Varnauskas,
H. B. Martin,
J. E. Cotes, and
R. L. Riley.
Relationship between cardiac output and apparent diffusing capacity of the lung in normal men during treadmill exercise.
J. Appl. Physiol.
13:
205-210,
1958.
|
| 229. |
Siebens, A. A.,
N. R. Frank,
D. C. Kent,
M. M. Newman,
R. A. Rauf, and
B. L. Vestal.
Measurements of the pulmonary diffusing capacity for oxygen during exercise.
Am. Rev. Respir. Dis.
80:
806-824,
1959.
|
| 230. |
Siegwart, B.,
P. Gehr,
J. Gil, and
E. R. Weibel.
Morphometry estimation of pulmonary diffusion capacity. IV. The normal dog lung.
Respir. Physiol.
13:
141-159,
1971.
|
| 231. |
Siggaard‐Andersen, O.
Blood acid-base alignment nomogram.
Scand. J. Clin. Lab. Invest.
15:
211-217,
1963.
|
| 232. |
Sjøgaard, G.
Electrolytes in slow and fast muscle fibers of humans at rest and with dynamic exercise.
Am. J. Physiol.
245
(Regulatory Integrative Comp. Physiol. 14):
R25-R31,
1933.
|
| 233. |
Sørensen, S. P. L.
Enzyme studien II. Uber die Messung und die bedeutung der Wasserstoffionen Koncentration bei enzymatischen Prozessen.
Biochem. Z.
21:
131-304,
1909.
|
| 234. |
Spriet, L. L.,
M. I. Lindinger,
R. S. McKelvie,
G. J. F. Heigenhauser, and
N. L. Jones.
Muscle glycogenolysis and H+ concentration during maximal intermittent cycling.
J. Appl. Physiol.
66:
8-13,
1989.
|
| 235. |
Spriet, L. L.,
C. G. Matsos,
S. J. Peters,
G. J. F. Heigenhauser, and
N. L. Jones.
Effects of acidosis on rat muscle metabolism and performance during heavy exercise.
Am. J. Physiol.
248
(Cell Physiol. 17):
C337-C347,
1985.
|
| 236. |
Spriet, L. L.,
K. Söderlund,
M. Bergstöm, and
E. Hultman.
Skeletal muscle glycogenolysis, glycolysis, and pH during electrical stimulation in men.
J. Appl. Physiol.
62:
616-621,
1987.
|
| 237. |
Stadie, W. C., and
H. O'brien.
The carbamate equilibrium II. The equilibrium between oxyhemoglobin and reduced hemoglobin.
J. Biol. Chem.
117:
439-470,
1937.
|
| 238. |
Stanley, W. C.,
E. W. Gertz,
J. A Wisnecki,
R. A. Neese,
D. L. Morris, and
G. A. Brooks.
Lactate extraction during net lactate release in legs of humans during exercise.
J Appl. Physiol.
60:
1116-1120,
1986.
|
| 239. |
Staub, N. C.,
J. M. Bishop, and
R. E. Forster.
Velocity of O2 uptake by human red cells.
J. Appl. Physiol.
17:
511-516,
1961.
|
| 240. |
Stewart, P. A.
Independent and dependent variables of acid-base control.
Respir. Physiol.
33:
9-26,
1978.
|
| 241. |
Stewart, P. A.
How to Understand Acid-Base. A Quantitative Acid-Base Primer for Biology and Medicine.
New York:
Elsevier/North Holland,
1981.
|
| 242. |
Stewart, P. A.
Modern quantitative acid-base chemistry.
Can. J. Physiol. Pharmacol.
61:
1444-1461,
1983.
|
| 243. |
Theorell, H.
Krisallinesches Myoglobin V. Mitteilung: Die Sauerstoffbindungskurve des Myoglobin.
Biochem. Z.
268:
73-82,
1934.
|
| 244. |
Thurlbeck, W. M.
The internal structure of nonemphysematous lungs.
Am. Rev Respir. Dis.
95:
765-773,
1967.
|
| 245. |
Triebwasser, J. H.,
R. L. Johnson, Jr.,
R. P. Burpo,
J. C. Campbell,
W. C. Reardon, and
C. G. Blomqvist.
Noninvasive determination of cardiac output by a modified acetylene rebreathing procedure utilizing mass spectrometer measurements.
Aviat. Space Environ. Med.
48:
203-209,
1977.
|
| 246. |
Trivedi, B., and
W. H. Danforth.
Effects of pH on the kinetics of frog muscle phosphofructokinase.
J. Biol. Chem.
241:
4110-4112,
1966.
|
| 247. |
Truog, W. E.,
M. P. Hlastala,
T. A. Standaert,
H. P. McKenna, and
W. A. Hodson.
Oxygen‐induced alteration of ventilation‐perfusion relationship in rats.
J. Appl. Physiol. Respir. Environ. Exerc. Physiol.
47:
1112-1117,
1979.
|
| 248. |
Turino, G. M.,
E. H. Bergofsky,
R. M. Goldring, and
A. P. Fishman.
Effect of exercise on pulmonary diffusing capacity.
J. Appl. Physiol.
18:
447-456,
1963.
|
| 249. |
van Leeuwen, A. M.
Net cation equivalency (“base binding power”) of the plasma proteins. A study of ion‐protein interaction in human plasma by means of in vivo ultrafiltration and equilibrium dialysis.
Acta Med. Scand.
176
(suppl. 422):
3-212, 176,
1964.
|
| 250. |
van Slyke, D. D.,
A. B. Hastings,
A. Hiller, and
J. Sendroy.
Studies of gas and electrolyte equilibria in blood. XIV. Amounts of alkali bound by serum albumin and globulin.
J. Biol. Chem.
79:
769-780,
1928.
|
| 251. |
Visser, B. F.
Pulmonary diffusion of CO2.
Phys. Med. Biol.
5:
155-166,
1960.
|
| 252. |
Wagner, P. D.,
B. K. Erikson,
K. Kubo,
A. Hiraga,
M. Kai,
Y. Yamaya,
R. S. Richardson, and
J. Seaman.
Maximum O2 transport and utilization before and after splenectomy.
In: Equine Exercise Physiology 4: Proceedings of the Fourth International Conference on Equine Exercise Physiology,
Korralbyn, Queensland, Australia:
Equine Vet. J.,
1994.
|
| 253. |
Wagner, P. D.,
G. E. Gale,
R. E. Moon,
J. R. Torre‐Bueno,
B. W. Stolp, and
H. A. Saltzman.
Pulmonary gas exchange in humans exercising at sea level and simulated altitude.
J. Appl. Physiol.
61:
260-270,
1986.
|
| 254. |
Wagner, P. D.,
J. R. Gillespie,
G. L. Landgren,
M. R. Feede,
B. W. Jones,
R. M. DeBowes,
R. L. Pieschl, and
H. H. Erickson.
Mechanism of exercise‐induced hypoxemia in horses.
J. Appl. Physiol.
66:
1227-1233,
1989.
|
| 255. |
Wagner, P. D.,
R. B. Laravuso,
R. R. Uhl, and
J. B. West.
Continuous distributions of ventilation‐perfusion ratios in normal subjects breathing air and 100% CO2.
J. Clin. Invest.
54:
54-68,
1974.
|
| 256. |
Wagner, P. D.,
H. A. Saltzman, and
J. B. West.
Measurements of continuous distributions of ventilation‐perfusion ratios: theory.
J. Appl. Physiol.
36:
588-599,
1974.
|
| 257. |
Wagner, P. D.,
J. R. Sutton,
J. T. Reeves,
A. Bymerman,
B. M. Groves, and
M. K. Malconian.
Operation Everest II: pulmonary gas exchange during a simulated ascent of Mt. Everest.
J. Appl. Physiol.
63:
2348-2359,
1987.
|
| 258. |
Wagner, P. D., and
J. B. West.
Effects of diffusion impairment on O2 and CO2 time courses in pulmonary capillaries.
J. Appl. Physiol.
33:
62-71,
1972.
|
| 259. |
Wagner, W. W., Jr., and
L. P. Latham.
Pulmonary capillary recruitment during airway hypoxia in the dog.
J. Appl. Physiol.
39:
900-905,
1975.
|
| 260. |
Wagner, W. W.,
L. P. Latham, and
R. L. Capen.
Capillary recruitment during airway hypoxia: role of pulmonary artery pressure.
J. Appl. Physiol.
47:
383-387,
1979.
|
| 261. |
Wang, C.‐W., and
A. S. Popel.
Effect of red bood cell shape on oxygen transport in capillaries.
Math. Biosci.
116:
89-110,
1992.
|
| 262. |
Wasserman, K.
The anaerobic threshold measurement to evaluate exercise performance.
Am. Rev. Respir. Dis.
129:
S35-S40,
1984.
|
| 263. |
Wasserman, K. W., and
B. J. Whipp.
Exercise physiology in health and disease.
Am. Rev. Respir. Dis.
112:
219-249,
1975.
|
| 264. |
Wiebel, E. R.
Morphometry of the Human Lung.
New York:
Academic Press, Inc.,
1963.
|
| 265. |
Weibel, E. R.
Morphometric estimation of pulmonary diffusion capacity. I. Model and method.
Respir. Physiol.
11:
54-75,
1970.
|
| 266. |
Weibel, E. R.
Morphometric estimation of pulmonary diffusion capacity. V. Comparative morphometry of alveolar lungs.
Respir Physiol.
14:
26-43,
1972.
|
| 267. |
Weibel, E. R.
A simpified morphometric method for estimating diffusing capacity in normal and emphysematous human lungs.
Am Rev. Respir Dis.
107:
579-588,
1973.
|
| 268. |
Weibel, E. R.
Morphometric and stereological methods in respiratory physiology, including fixation techniques. Techniques in the life sciences.
Respir. Physiol.
P401:
1-35,
1984.
|
| 269. |
Weibel, E. R.,
W. J. Federspiel,
F. Frydor‐Doffey,
C. C. W. Hsia,
M. Konig,
V. Stalder‐Navarro, and
R. Vock.
Morphometric model for pulmonary diffusing capacity I. Membrane diffusing capacity.
Respir. Physiol.
93:
125-149,
1993.
|
| 270. |
Weibel, E. R., and
B. W. Knight.
A morphometric study on the thickness of the pulmonary air‐blood barrier.
J. Cell Biol.
21:
367-384,
1964.
|
| 271. |
Weibel, E. R.,
L. B. Marques,
M. Constantinopol,
F. Doffey,
P. Gehr, and
C. R. Taylor.
Adaptive variation in the mammalian respiratory system in relation to energetic demand: VI. The pulmonary gas exchanger.
Respir. Physiol.
69:
81-100,
1987.
|
| 272. |
Weibel, E. R.,
C. R. Tayor,
J. J. O'Neil,
D. E. Leith,
P. Gehr,
H. Hoppeler,
V. Langman, and
R. V. Baudinette.
Maximal oxygen consumption and pulmonary diffusing capacity: a direct comparison of physiologic and morphometric measurements in canids.
Respir. Physiol.
54:
173-188,
1983.
|
| 273. |
Weibel, E. R.,
P. Untersee,
J. Gil, and
M. Zulauf.
Morphometric estimation of pulmonary diffusion capacity. VI. Effect of varying positive pressure inflation of air spaces
Respir. Physiol.
18:
285-308,
1973.
|
| 274. |
West, J. B.,
K. Tsukimoto,
O. Matthieu‐Costello, and
R. Predilleto.
Stress failure in pulmonary capillaries.
J. Appl. Physiol.
70:
1731-1742,
1991.
|
| 275. |
West, J. B., and
P. D. Wagner.
Pulmonary gas exchange.
In: Bioengineering Aspects of the Lung,
edited by J. B. West.
New York and Basel:
Marcel Dekker, Inc.,
1980.
|
| 276. |
Whipp, B. J.
The bioenergetic and gas exchange basis of exercise testing.
Clin. Chest Med.
15:
173-192,
1994.
|
| 277. |
Whipp, B. J.,
J. A. Davis, and
K. Wasserman.
Ventilatory control of the “isocapnic buffering” region in rapidly‐incremental exercise.
Respir. Physiol.
76:
357-368,
1989.
|
| 278. |
Whipp, B. J., and
S. A. Ward.
Coupling of ventilation to pulmonary gas exchange during exercise.
In: Exercise. Pulmonary Physiology and Pathophysiology,
edited by B. J. Whipp and
K. Wasserman.
New York:
Marcel Dekker, Inc.
1991,
p. 271-307.
|
| 279. |
Wilhelm, E.,
R. Baiting, and
J. Wilcock.
Low pressure solubility of liquid gases in water.
Chem. Rev.
77:
219-262,
1977.
|
| 280. |
Wittenberg, J. B.
Myogobin‐facilitated oxygen diffusion: role of myoglobin in oxygen entry into muscles.
Physiol. Rev.
50:
559-636,
1970.
|
| 281. |
Worth, H.,
W. Nusse, and
J. Piiper.
Determination of binary diffusion coefficients of various gas species used in respiratory physiology.
Respir. Physiol.
32:
15-26,
1978.
|
| 282. |
Worth, H., and
J. Piiper.
Diffusion of helium, carbon monoxide and sulfur hexafluoride in gas mixtures similar to alveolar gas.
Respir. Physiol.
32:
155-166,
1978.
|
| 283. |
Wranne, B.,
R. D. Woodson, and
J. C. Detter.
Bohr effect: interaction between H+, CO2, and 2, 3 DPG in fresh and stored blood.
J. Appl. Physiol.
32:
749-754,
1972.
|
| 284. |
Wyman, J., Jr.
Heme proteins,
Adv. Protein Chem.
4:
407-531,
1948.
|
| 285. |
Wyman, J.
Facilitated diffusion and the possible role of myoglobin as a transport mechanism.
J. Biol. Chem.
241:
115-121,
1966.
|
| 286. |
Yamaguchi, K.,
P. D. Nguyen,
P. Scheid, and
J. Piiper.
Kinetics of O2 uptake and release by human erythrocytes studied by a stopped‐flow technique.
J. Appl. Physiol.
58:
1215-1224,
1985.
|
| 287. |
Yamamoto, W. S., and
M. W. Edwards, Jr..
Homeostatis of carbon dioxide during intravenous infusion of carbon dioxide.
J. Appl. Physiol.
15:
807-818,
1960.
|
| 288. |
Yoshida, F., and
N. Oshima.
Diffusivity of oxygen in blood serum.
J. Appl. Physiol.
21:
915-919,
1966.
|